Nesneleri tanıma yeteneği, görsel hayvanların hayatta kalması için temeldir. Primatların ventral akışı uzun süredir nesnelerin beyinde nasıl işlendiğini incelemek için bir model olarak hizmet ediyor10,11. Primat ventral akışının tanımlayıcı özelliklerinden biri hiyerarşik organizasyondur.12Nesne tanıma konusunda eğitilmiş derin sinir ağları (DNN’ler) tarafından yansıtılan8,13. Bu paralellik önemli bir soruyu gündeme getiriyor: Hiyerarşik temsil gerekli midir ve eğer öyleyse, görselliği yüksek olan tüm memeli türlerinde bulunabilir mi? Farklı memeli türlerinde görsel işlemenin araştırılması, nesne görüşüne ilişkin genel prensiplerin daha derin anlaşılmasını sağlamayı vaat ediyor.
On yıldan fazla bir süre önce, fare sinir devresi diseksiyonu için mevcut araçların zenginliği nedeniyle, fare görsel sistemi büyük ilgi görmeye başladı.14,15. Ancak farenin düşük görme keskinliği ve görmeye ayrılmış kortikal bölgesinin sınırlı olması16 nesne tanımanın altında yatan hiyerarşik beyin mekanizmalarını incelemek için onu ideal olmayan bir organizma haline getiriyor. Ağaç sivri faresi, görsel işlemeyi incelemek için bir model olarak giderek artan ilgi görüyor17 Yüksek görme keskinliği sayesinde (kemirgenlerin on katından fazla)18büyük ölçüde genişletilmiş görsel korteks19 ve fareyle karşılaştırıldığında görsel olarak yönlendirilen davranışsal görevleri yerine getirme konusunda mükemmel yetenek20,21. Ağaç sivri faresi görsel sistemi en az dokuz farklı anatomik görsel kortikal alan içerir19. Birincil görsel alan (V1), yönlendirme seçici sütunların düzenli bir şekilde düzenlenmesi de dahil olmak üzere yüksek derecede işlevsel uzmanlaşma gösterir.22,23. Ağaç sivri faresi ayrıca, primatlardan farklı olan geniş ölçekli bir topografik organizasyona sahip olmasına rağmen, belirgin bir ikinci görsel alana (V2) sahiptir.24. Lezyon çalışmaları, V2’nin önündeki ağaç faresi ekstrastriat alanları ile primat IT korteksi arasında kaba bir benzerlik olduğunu göstermektedir: temporal lobun büyük bölümlerinin ablasyonları, primatlardaki alt temporal (IT) lezyonların etkilerine benzer şekilde desen ayrımcılığı ve nesne görüşünde eksiklikler üretir.19,25,26. Ancak bizim bilgimize göre, ağaç faresinin V2’nin ötesindeki ekstrastriate görsel alanlarının fonksiyonel özelliklerine ilişkin elektrofizyolojik çalışmalar yapılmamıştır.
Burada, tüm ağaç faresi ventral akışı boyunca görsel nesne temsilinin altında yatan kortikal organizasyonu ve kodlama ilkelerini tanımlamayı hedefliyoruz. Birkaç Neuropixels sondası ile büyük ölçekli elektrofizyolojik kayıtları kullanarak, pulvinarın yanı sıra beş ağaç faresinin ventral görsel alanını araştırdık. Artan alıcı alan boyutu ve yanıt gecikmesi dahil olmak üzere primatlarda bulunan hiyerarşik organizasyonun ayırt edici özelliklerini doğruladık.27 spektral olarak eşleşen gürültüyle karşılaştırıldığında doğal dokular için artan seçiciliğin yanı sıra5. Ağaç faresindeki V2 alanının, primat BT korteksiyle ilişkili temel işlevleri yerine getirdiğini bulduk. Bu, yüksek seviyeli nesne alanının tam temsilini, doğru nesne kimliğinin kodunun çözülmesini ve yeniden yapılandırılmasını ve güçlü yüz seçici hücrelerin varlığını içerir. Genel olarak sonuçlar, primatlarda daha önce gözlemlenen temsillerin görsel işlemenin çok daha erken bir aşamasında gerçekleştiği, ağaç faresinde sıkıştırılmış, çok aşamalı bir hiyerarşiye işaret ediyor.
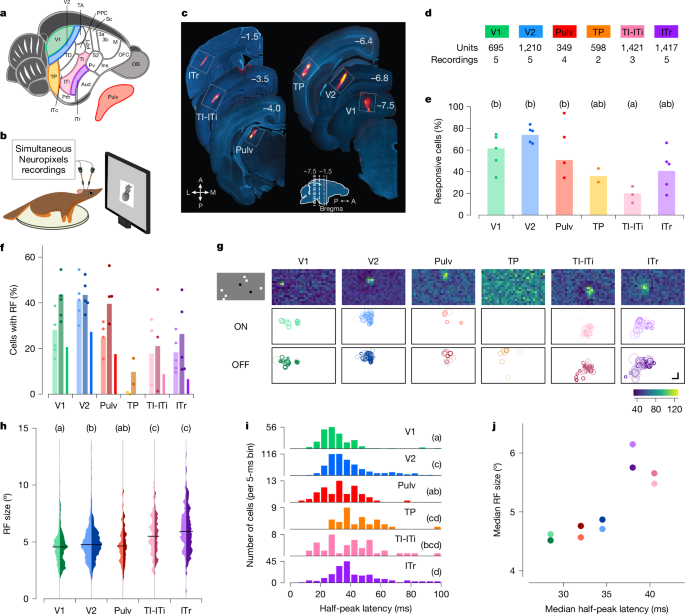
Hiyerarşik görsel işlemeyi araştırmak için ağaç faresinin ventral akışını kapsayan bir dizi alanı hedefledik (Şekil 1). 1a). Birincil (V1) ve ikincil (V2) görsel alanları, görsel işlemenin erken aşamalarında yer alan mimari açıdan farklı bölgeler olarak dahil ettik.28,29. Ventral görsel işleme akışı boyunca potansiyel bir ara düğüm olarak temporal posterior (TP) alanı seçtik. Ön uçta, makak IT korteksine homolog olabilecek üç alt bölgeye odaklandık: temporal-inferior (TI), temporal orta (ITi) ve inferotemporal rostral (ITr) alanlar. TI ve ITi lezyonları görsel form tespitinde ciddi bozulmalara neden olur25. ITr hem görsel hem de işitsel korteksten girdi alır19ancak görsel işlevsel özellikleri bildiğimiz kadarıyla hiç araştırılmamıştır. TI ve ITi arasındaki sınırın ayırt edilmesindeki zorluk nedeniyle bunları bir arada grupladık ve bu bölgeyi TI-ITi olarak adlandırdık. Çünkü birçok temporal bölge talamustan doğrudan girdi alır.30pulvinarın (Pulv) dorsal görsel kısmını da kayıtlarımıza dahil ettik. Elektrot hedeflemeyi yönlendirmek için geriye dönük izleme deneyleri yaptık (Genişletilmiş Veri Şekil 1). 1a,b).

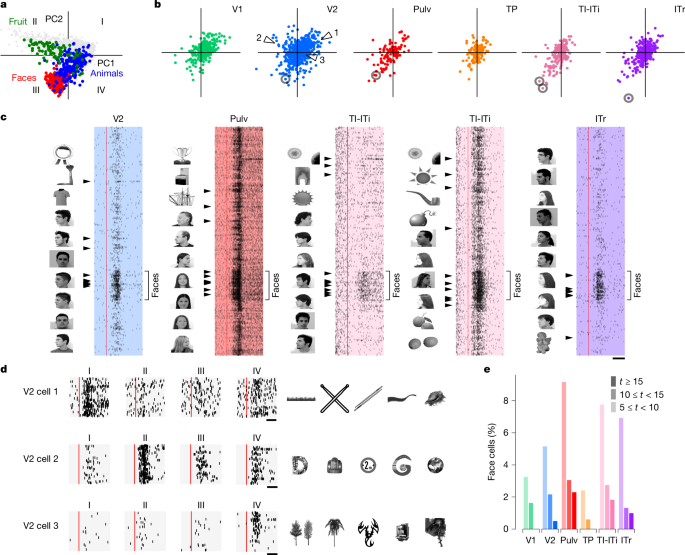
V1, V2, TP, TI-ITi, ITr ve Pulv’daki nöronların görsel tepkilerini karakterize etmek için uyanık ağaç farelerinde Neuropixels problarını kullanarak elektrofizyolojik kayıtlar yaptık (Şekil 1). 1b). Her deney sırasında, hayvanların kafası bir monitörün önüne sabitlendi ve yerel seyrek gürültü, statik ızgaralar, doğal dokular ve gürültü ile yüzlerin ve diğer nesnelerin görüntüleri dahil olmak üzere bir dizi görsel uyaran sunuldu. Her oturumun sonunda prob konumları DiI (DiIC18(3), bir floresan boya) ile işaretlendi ve hedefleme histoloji ile doğrulandı (Şekil 1). 1c). Test ettiğimiz görsel uyaran sınıflarından herhangi birine yanıt veren bir hücreyi görsel olarak duyarlı olarak sınıflandırdık (Yöntemler). Her alanda çok sayıda iyi izole edilmiş tekil ünite bulduk (Şekil 1). 1d), görsel uyaranlara yanıt veren hücrelerin fraksiyonlarında bazı alanlar arası farklılıklarla (ANOVA, F5,18 = 5,362, P= 0,003; İncir. 1e). Özellikle, V2 hücreleriyle karşılaştırıldığında önemli ölçüde daha az sayıda TI-ITi hücresi görsel olarak duyarlıydı.
Yerel olarak seyrek bir gürültü uyaranı kullanarak ağaç sivri faresinin ventral yolu boyunca nöronların alıcı alanlarını haritalayarak başladık (Yöntemler). Her bir nöron için, görme alanı konumlarındaki ani artış sayımlarının iki boyutlu (2D) matrisine bir Gauss dağılımını uydurarak alıcı alanı tahmin ettik; AÇIK ve KAPALI alıcı alanları, sırasıyla beyaz ve siyah karelere verilen yanıtlar kullanılarak ayrı ayrı hesaplandı. AÇIK ve / veya KAPALI alıcı alanlara sahip hücreler, TP dışındaki tüm alanlarda açıkça mevcuttu (Şekil 1). 1f). Bu, en öndeki iki alan olan TI-ITi ve ITr’yi içeriyordu; bu, nöronların tipik olarak mekansal olarak değişmez tepkiler gösterdiği primatlardaki ön temporal lobla çelişir.31,32.
Bireysel kayıtlarda, alıcı alan pozisyonları, elektrotla hedeflenen kortikal bölge tarafından temsil edilen retinotopik bölgeye karşılık gelen, görme alanının küçük bir bölümünde kümelenmiştir. Figür 1g her alan için temsili bir oturumda kayıtlı tüm hücrelerin alıcı alanlarını gösterir. Bu kümelenme, en öndeki alanlar olan TI-ITi ve ITr dahil olmak üzere incelenen tüm alanlarda belirgindi. Bu bulgu, ventral akımın ön ucundaki konumlarına rağmen bu alanların retinotropik organizasyonu koruduğunu göstermektedir.
Kaydedilen alanlar arasındaki hiyerarşik ilişkileri değerlendirmek için öncelikle hiyerarşik düzeyin iki klasik ölçümünü inceledik: alıcı alan boyutu ve görsel olarak uyarılmış yanıt gecikmesi. Alıcı alan boyutları sistematik olarak arkadan öne doğru arttı (Şekil 1). 1 saat). Ayrıca her bölgedeki her ünite için yarı tepe gecikmelerini hesapladık ve gecikmelerin V1’den V2’ye ve ITr’ye arttığını bulduk (Şekil 1). 1i Ve Yöntemler). Alıcı alan boyutlarındaki artışın öngördüğü hiyerarşi, gecikmelerdeki artışın öngördüğü hiyerarşiyle genel olarak tutarlıydı (Şekil 1). 1 gün).
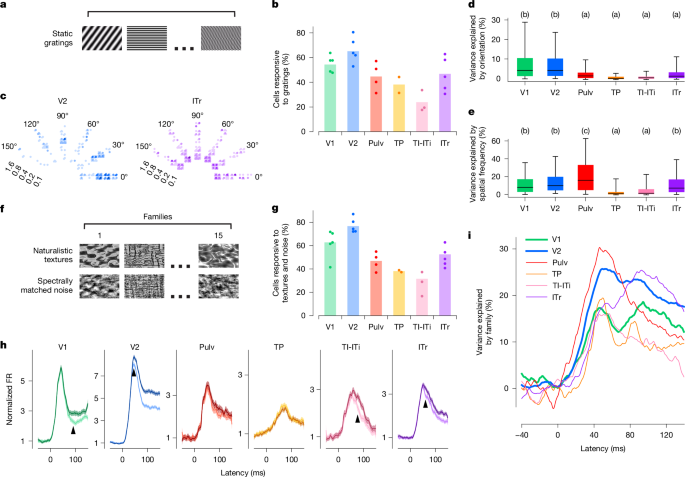
Primat görsel kortekste erken görsel alanlar, yönelim ve uzamsal frekans gibi düşük seviyeli özelliklere güçlü bir şekilde ayarlanırken, sonraki alanlar daha karmaşık nesne özelliklerine göre ayarlanır.7,33,34,35. Ağaç sivri faresinde benzer bir ilerlemenin olup olmadığını incelemek için, statik ızgaralar kullanarak ventral görsel alanlar boyunca yönelimi ve uzaysal frekansı ayarlamayı değerlendirdik (Şekil 1). 2a). Görsel olarak duyarlı nöronların oranının (bkz. 1eIzgaralara yanıt veren V1 ve V2’de en yüksek (sırasıyla yaklaşık %55 ve %65) ve TI-ITi’de en düşüktü (Şekil 1). 2b). V2 ve ITr’deki örnek hücrelerin oryantasyonuna, uzamsal frekansına ve fazına ayarlama, ağaç sivri faresi görsel alanlarında bu değişkenlere gözlemlediğimiz çeşitli ayarlamaları göstermektedir (Şekil 1). 2c). Genel olarak, oryantasyon ayarlaması en çok V1 ve V2’de yaygındı (ANOVA’dan sonra Tukey analizi, F5, 1,106 = 26,791, P< 10−24İncir. 2 gün), oysa uzaysal frekans ayarı ITr’de de yaygındı (Tukey analizi ANOVA, F5, 1106 = 20.514, P< 10−18İncir. 2e). Bu bulgular, oryantasyon ayarının özellikle erken görsel alanlarda belirgin olduğu ve daha sonraki alanlarda keskin bir şekilde azaldığı primat ve kemirgenlerin ventral akışında bulunanlarla kabaca tutarlıdır.36,37,38,39.
Şu ana kadar V2 yanıtları büyük ölçüde V1’dekilere benzer görünüyordu ve bu da V2’nin herhangi bir farklı hesaplama işlevi gerçekleştirip gerçekleştirmediği sorusunu gündeme getirdi. Makaklarda, doğal dokulardaki yüksek dereceli istatistiksel bağımlılıklara duyarlılık, V2 alanının ayırt edici bir özelliği olarak tanımlanmıştır (ref. 5). Bu nedenle, ağaç farelerinin ekstrastriat alanlarının doğal doku işleme konusunda benzer bir uzmanlık gösterip göstermediğini sorduk. Bunu test etmek için, doğal dokular ve spektral olarak eşleşen sentetik gürültü görüntüleri sunarken altı görsel alanın tamamında sinirsel aktivite kaydettik (Şekil 1). 2f Ve Yöntemler). Tüm alanlar arasında V2, doku ve/veya gürültü uyaranlarına yanıt veren en yüksek hücre oranını içeriyordu (Şekil 1). 2g). Popülasyon tepki dinamikleri, V2’deki doğal dokular ve gürültü arasındaki en güçlü diferansiyel aktiviteyi ortaya çıkardı, bunu V1, ITr ve TI-ITi takip etti ve geri kalan alanlarda minimum modülasyon veya hiç modülasyon olmadı (Şekil 1). 2 saat). V2’de fark, uyaranın süresi boyunca devam etti. Her ne kadar V1’deki yanıtlar V2’dekilerden çok daha önce başlamış olsa da (Şekil 1). 1i), doku ve gürültü tepkileri arasındaki farklılık V1’de (90 ms’de) V2’ye (45 ms’de) göre daha sonra meydana geldi; bu, V1’deki doku modülasyonunun V2’den gelen geri bildirim yoluyla ortaya çıkabileceğini gösteriyor. Bu yorum, V2’nin doku ailesi kimliğini V1’den daha önce kodladığının bulunmasıyla da desteklenmektedir (Şekil 1). 2i).
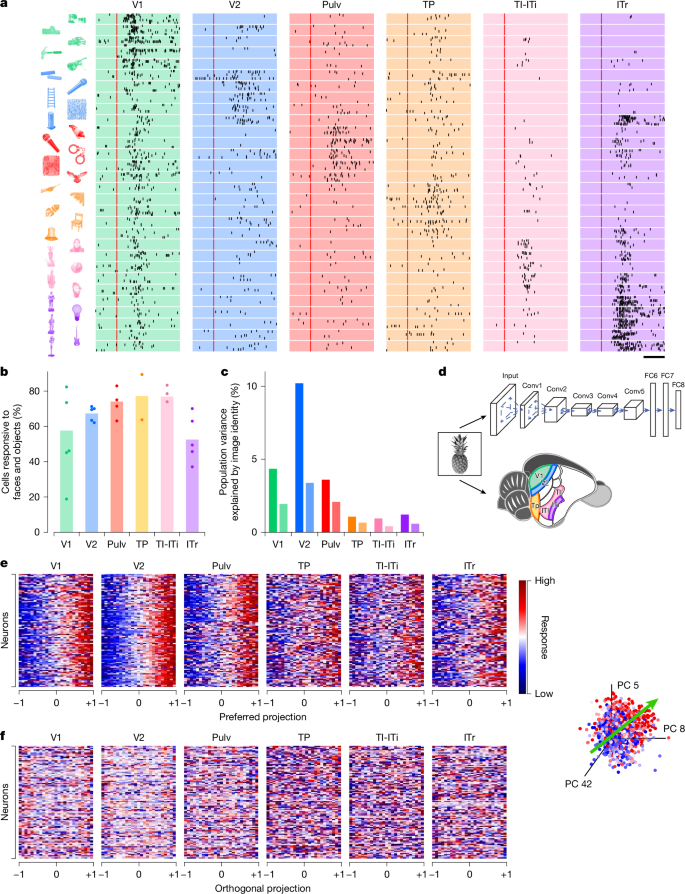
Görsel hiyerarşinin merkezi bir işlevi, navigasyon, yiyecek arama veya çiftleşme gibi hayati davranışlara rehberlik edecek nesneleri tanımak ve sınıflandırmaktır. Ağaç faresinin ventral akışındaki yüksek seviyeli nesne temsillerini araştırmak için hayvanların, vücut parçalarının, yüzlerin ve gündelik nesnelerin 1.593 görüntüsünden oluşan zengin bir uyaran seti sunduk (Yöntemler). Aynı uyaran seti daha önce makak alt temporal (IT) korteksindeki ayarlamayı karakterize etmek için kullanılmış ve primatlarda ve ağaç farelerinde nesne tanıma mekanizmaları arasında doğrudan karşılaştırmalar yapılmasına olanak sağlanmıştır.8. Uyaranlar, kaydedilen nöronların alıcı alan konumuyla eşleşecek şekilde ayarlandı (Yöntemler). Örnek hücrelerden alınan yanıt rasterleri, ağaç faresinin ventral akışındaki farklı nöronlar arasında nesne seçiciliğinde çeşitlilik gösterdi (Şekil 1). 3a). Kaydedilen altı alan arasında görsel olarak duyarlı hücrelerin benzer bir oranı, V2, TP, TI-ITi ve Pulv’daki nesne uyaranlarına yanıt verdi (Şekil 1). 3b). Özellikle, TI-ITi’deki görsel olarak duyarlı hücrelerin çok daha büyük bir kısmı, ızgaralara kıyasla nesne uyaranlarına yanıt verdi (Şekil 1). 2b), görsel hiyerarşide daha yüksek bir seviyeyi işgal eden zamansal alanlarla tutarlıdır. Nesneye dayalı yanıtların güvenilirliğini ölçmek için, ‘açıklanabilir varyansı’ (nöral yanıt varyansının denemeden denemeye değişkenlik yerine uyaran kimliğine atfedilebilen kısmı) hesapladık (Yöntemler). V2’den sonra, bu karmaşık nesne uyaranlarına verilen yanıtlardaki açıklanabilir değişkenlik önemli ölçüde azaldı (Şekil 1). 3c), daha ön bölgelerdeki yanıtların denemeler arasında daha az tutarlı olduğunu gösteriyor. Açıklanabilir varyansın düşük seviyeli görsel özelliklerle açıklanıp açıklanamayacağını belirlemek için parlaklık, kontrast ve uzaysal frekansın katkılarını analiz ettik; her alanda, varyansın yalnızca küçük bir kısmı bu özelliklerle açıklanabilmektedir (Şekil 1). 3c ve Genişletilmiş Veri Şek. 2).
Her alanın kullandığı sinir kodunun doğasını daha iyi anlamak için sinir tepkilerini AlexNet’i kullanarak modelledik.40nesne tanıma konusunda eğitilmiş sekiz katmanlı bir DNN (Şekil 1). 3 boyutlu). Makaklarda, tek BT nöronları, her hücrenin gelen uyaranları DNN’den türetilmiş bir özellik alanında tercih edilen bir eksene doğrusal olarak yansıttığı bir ‘eksen modeli’ ile iyi bir şekilde tanımlanır.8,13. Bu modellerde tercih edilen eksenler nispeten düşük boyutlu bir temele yayılır; örneğin makak yüz parçalarından yüzlerin doğru şekilde yeniden yapılandırılması için yalnızca 50 boyut yeterlidir.41. Bu prensibin ağaç faresinde de geçerli olup olmadığını test etmek için, AlexNet FC6 katmanındaki ilk 50 temel bileşeni kullanarak her bir nöronun tercih edilen eksenini altı kayıtlı alan boyunca hesapladık. Ağaç sivri faresi korteksinin, makak BT korteksinde gözlemlendiği gibi, yüksek seviyeli bir nesne alanını temsil edip etmediğini açıklığa kavuşturmak için FC6’ya odaklandık.8. Eksen bazlı kodlamayla tutarlı olarak, altı alanın tamamındaki nöronlar, tercih ettikleri eksenler boyunca rampa şeklinde ayarlama gösterdi (Şekil 1). 3e Ve Yöntemler). Dahası, hücreler ana ortogonal eksenleri boyunca düz bir ayar gösterdi (yani, tercih edilen eksene dik olan en uzun eksen; Şekil 1). 3f Ve Yöntemler).
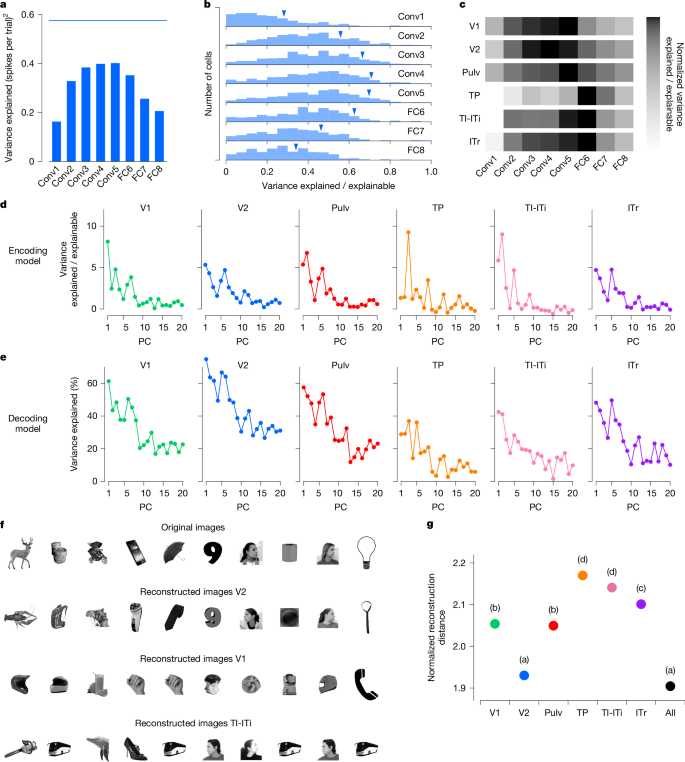
Primatlarda yapılan önceki çalışmalar, AlexNet’in ve diğer DNN’lerin erken katmanlarının, erken retinotopik görsel alanlardaki nöronal aktiviteyi en iyi şekilde açıkladığını, geç katmanların ise IT korteksteki yanıtları en iyi şekilde açıkladığını göstermiştir.8,13. Ağacın sivri faresinin karın akıntısında da benzer bir modelin geçerli olup olmadığını sorduk. Bunu test etmek için, AlexNet’teki her katmanın ilk 50 temel bileşenine göre tek hücreli ateşleme oranlarını regrese ettik (Yöntemler) ve her hücrenin tepkisindeki varyansı en iyi açıklayan katmanı belirledi. V2’deki temsili bir hücre için AlexNet katmanı Conv4, yanıtlarını en iyi şekilde açıkladı (Şekil 1). 4a). V2 popülasyonunda, ara katmanların (özellikle Conv4 ve Conv5) tutarlı bir şekilde erken veya geç katmanlardan daha fazla açıklayıcı güce sahip olduğunu bulduk (Şekil 1). 4b).
Farklı AlexNet katmanlarının beyin alanları arasındaki açıklayıcı gücünü karşılaştırmak için, çeşitli AlexNet katmanları tarafından açıklanan varyansın her bir alanı içindeki hücreler arasındaki toplamı hesapladık ve bu toplamları, açıklanabilir varyanslarının hücreler arası toplamı ile normalleştirdik (Yöntemler). Bu analiz, erken görsel alanlar V1 ve V2’nin en iyi şekilde ara katmanlar (özellikle Conv3’ten Conv5’e) tarafından açıklandığını, oysa ön alanlar TI-ITi ve ITr’nin en iyi şekilde yüksek seviyeli FC6 katmanı tarafından açıklandığını ortaya çıkardı (Şekil 1). 4c). Bununla birlikte, AlexNet tarafından açıklanan mutlak varyans, bu yüksek kortikal alanlarda daha düşüktü (Genişletilmiş Veri Şekil 1). 3a,b), ön bölgelerde gözlemlenen nesne kimliğine verilen yanıtların denemeden denemeye güvenilirliğinin azalmasıyla tutarlıdır (Şekil 1). 3c). Olası bir açıklama, AlexNet’in, multimodal olduğu ve yalnızca görsel olmadığı öne sürülen ön ağaç faresi bölgelerinin tepki özelliklerini tam olarak yakalamak için ifade kapasitesinden yoksun olabileceğidir.19
Hangi özellik eksenlerinin alanlar arasındaki nöral tepkilerde en fazla varyansı açıkladığını araştırmak için, AlexNet FC6 katmanındaki bireysel özellik temel bileşenleri tarafından ne kadar varyansın açıklandığını inceledik. Genel olarak, daha önceki temel bileşenler, alanlar arasında bazı değişkenliklerle birlikte, nöral tepkilerdeki varyansın en büyük oranını açıkladı (Şekil 1). 4 gün). Ayrıca, her bir görsel alandaki popülasyon aktivitesinden spesifik FC6 özelliklerinin ne kadar iyi çözülebileceğini de analiz ettik (Şekil 1). 4e). Yine, ilk temel bileşenler en güçlü şekilde temsil edildi; kod çözme performansı V2’de zirveye ulaştı ve diğer tüm bölgelere göre önemli ölçüde daha yüksekti. Bu bulgu, FC6 özelliklerinin V2’de diğer alanlara göre daha fazla varyansı açıkladığı gözlemiyle uyumludur (Genişletilmiş Veri Şekil 1). 3c). Bu nedenle, V2 en iyi şekilde Conv4 ve Conv5 özellikleriyle açıklansa da, TI-ITi ve ITr en iyi şekilde FC6 özellikleriyle açıklansa da FC6 özellikleri yine de V2’de bu daha ön bölgelere göre daha iyi temsil edildi.
V2’nin AlexNet FC6 özelliklerinin kodunu çözmedeki güçlü performansı göz önüne alındığında, daha önce maymun IT korteksinde gösterildiği gibi, V2’deki aktivitenin küçük sinir popülasyonları kullanarak nesneleri yeniden yapılandırmak için yeterli olup olmayacağını sorduk.8. Bunu test etmek için, her biri FC6 aktivasyonlarını çıkarmak için AlexNet’ten geçen 15.901 görüntüden oluşan büyük bir yardımcı veri kümesi kullandık. Her alandaki aktiviteden bir FC6 aktivasyon vektörünü yeniden oluşturduk ve FC6 özellikleri yeniden yapılanmaya en yakın olan görüntüyü belirledik (Genişletilmiş Veri Şekil 1). 3 boyutlu). Hücre sayısını kontrol etmek için her alandan rastgele seçilen 100 hücreyi kullanarak yeniden yapılandırmalar gerçekleştirdik. Parametre kodu çözme konusundaki sonuçlarımızla tutarlıdır (Şekil 1). 4eV2’de optimal olan V2’den yeniden oluşturulan görüntüler orijinal görüntülere çok benziyordu, oysa V1 veya TI-ITi’den yeniden oluşturulan görüntüler belirgin şekilde daha az doğruydu (Şekil 1). 4f). Yeniden yapılandırma doğruluğunu alanlar arasında niceliksel olarak karşılaştırmak için, her görüntü için yeniden yapılandırılmış ve gerçek FC6 aktivasyon vektörleri arasındaki mesafeyi hesapladık ve teorik en iyi kod çözme mesafesiyle normalleştirildi (Yöntemler). Bu analiz, V2’nin tüm alanlar arasında en küçük normalleştirilmiş kod çözme mesafelerine sahip olduğunu (en doğru yeniden yapılandırmaları gösterir) ve tüm alanlardaki nöronların bir araya toplanmasıyla elde edilen performansla eşleştiğini ortaya çıkardı (ANOVA’dan sonra Tukey analizi, F6, 11144= 151.248, P< 10−184; İncir. 4g). Bu sonuçlar ayrıca ağaç faresi V2’de bulunan zengin ancak kompakt nesne temsilinin altını çizmektedir.
Primat BT korteksi, yüzler gibi belirli kategorilerdeki görüntülere maksimum düzeyde yanıt veren nöronlardan oluşan bölgeleri içerir.42,43,44. Bu tür kategori seçici bölgeler, BT korteksinin genel bir nesne alanını (AlexNet FC6 özelliklerinin ilk iki ana bileşeni tarafından tanımlanan bir temsil alanı) kodladığı normatif bir çerçeve ile açıklanabilir.8,45. Bu alanda farklı sektörler yüzler, meyveler ve hayvanlar gibi farklı nesne kategorilerine karşılık gelir (Şekil 1). 5a).
Ağacın görsel korteksi, primat BT korteksi gibi, nesne alanının farklı sektörlerini temsil etmek için uzmanlaşmış bölgeler içeriyor mu? Bu soruyu yanıtlamak için, kaydedilen tüm hücrelerin tercih edilen eksenlerini aynı 2 boyutlu nesne alanına yansıttık (Şekil 1). 5b). V2’de tercih edilen eksenler dört çeyreğin tamamına dağıtılırken diğer alanlarda büyük ölçüde I ve III. çeyreklerle sınırlıydı. Farklı nesne kategorilerinin bu alanın farklı bölgelerine lokalize olduğu göz önüne alındığında, bireysel ağaç faresi nöronlarının belirli kategoriler için seçicilik göstereceğini tahmin ettik. Aslında, yanıt rasterlerinin analizi, yüz sektöründe tercih edilen eksenlere sahip nöronların güçlü bir şekilde yüz seçici olduğunu doğruladı (Şekil 1). 5c). Bazı yüz hücreleri aynı zamanda diğer yuvarlak şekillere de tepki verirken, diğerleri yalnızca yüzler için güçlü seçicilik gösterdi. Yüz hücrelerine ek olarak, dikenli, uzun nesneler (çeyrek I), yuvarlak cansız nesneler (bölge II) ve dikenli hareketli nesneler (çeyrek IV) için seçici nöronlar belirledik (Şekil 1). 5 gün ve Genişletilmiş Veri Şek. 4a,b). Bununla birlikte, primat BT’de görülen modüler organizasyonun aksine, ağaç farelerinin görsel alanlarında kategori seçici nöronların topografik kümelenmesine dair hiçbir kanıt bulamadık (Genişletilmiş Veri Şekil 1). 4c).
Yüzlerin, özellikle de tüm yüz uyaranlarımızı oluşturan insan yüzlerinin, ağaç fareleri için özel bir davranışsal öneme sahip olduğu bilinmemektedir.46. Hücrelerin gerçekten yüz seçici olduğunu doğrulamak için, her bir hücre için yüzlere ve diğer tüm nesnelere verilen tepkiler arasındaki fark olarak tanımlanan bir yüz seçicilik indeksi hesapladık (Yöntemler). Bu, yüksek derecede yüz seçici hücrelerin küçük popülasyonlarını doğruladı (T≥ 15) çoğu alanda V2 alanından başlayarak, en yüksek yüzdeler TI-ITi ve Pulv’dadır (Şekil 1). 5e).
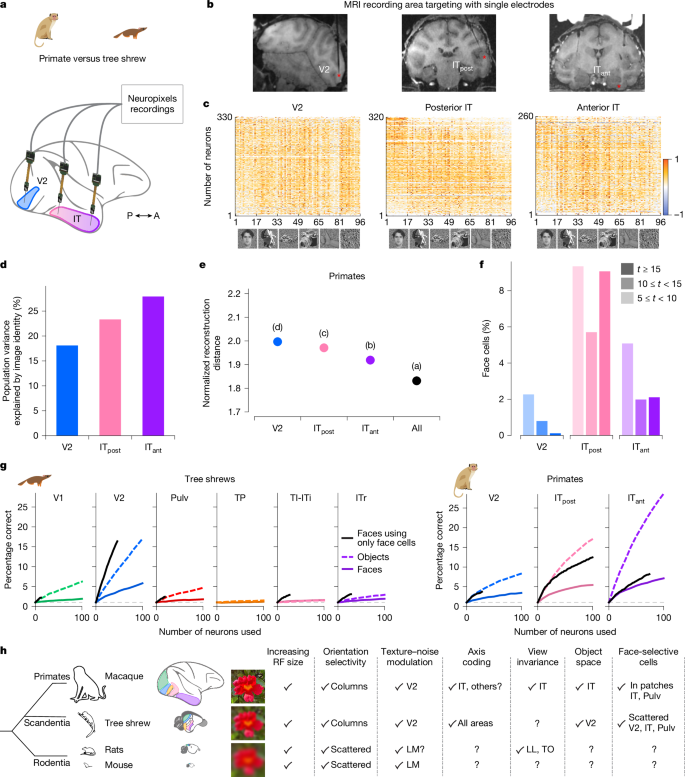
Primate IT korteksi, nesne tanıma konusunda oldukça uzmanlaşmıştır ve uzun süredir görsel form işlemenin incelenmesi için bir temel olarak hizmet etmektedir. Ağaç faresi veri setimizle doğrudan karşılaştırmalar yapabilmek için NHP Neuropixels problarını kullanarak makak maymunlarında büyük ölçekli kayıtlar gerçekleştirdik. Aynı 1.593 nesne uyaranını V2, arka IT’den (IT) kaydederken sunduk.postalamak) ve ön BT (BTkarınca) alan başına iki maymundan (Şek. 6a-c). Karmaşık nesne uyaranlarına verilen yanıtlardaki açıklanabilir varyansın, primat V2’den IT’ye kadar primat görsel hiyerarşisi boyunca arttığını bulduk.karınca(İncir. 6 gün), oysa ağaç faresinin görsel korteksinde V2’de zirveye ulaştı (Şekil 1). 3c). Benzer şekilde, görüntü yeniden yapılandırma performansı primat hiyerarşisi boyunca gelişti (Şekil 1). 4g), oysa ağaç farelerinde bu en doğru V2’deydi (Şekil 1). 6e). Ağaç sivri farelerinin aksine (Şek. 5e), primat V2’de güçlü bir şekilde yüz seçici hücreler gözlemlemedik (Şekil 1). 6f). Beklendiği gibi primat BT’deki yüz hücrelerinin sayısıpostalamakve BTkarıncaçok daha yüksekti. Özellikle, BT’lerden birindepostalamakKayıtlarda, prob kısmen bilinen bir yüz parçasını hedef aldı ve bu da daha yüksek oranda yüz hücresi oluşmasına neden oldu.
Son olarak, primat ve ağaç sivri faresi görsel sistemlerindeki sinir popülasyonlarının bireysel yüz veya nesne kimliğini ne kadar iyi çözebildiğini sorduk. Bunu test etmek için sınıflandırıcıları, her alandaki rastgele örneklenmiş alt popülasyonlardan alınan sinirsel aktiviteyi kullanarak 100 yüzün veya 100 genel nesnenin kimliğini çözecek şekilde eğittik (Şekil 1). 6g Ve Yöntemler). Ağaç farelerinde, TP dışındaki tüm alanlar, hem yüzler hem de nesneler için şansın üzerinde kod çözme performansı gösterdi. Analizi yalnızca yüz seçici hücrelerle sınırladığımızda yüz kimliğinin kod çözümü daha da gelişti. Ağaç faresi V2’deki kod çözme performansı, hem yüz hem de nesne kimliği açısından diğer tüm ağaç faresi alanlarındaki performansı aştı. Buna karşılık, primat V2, ağaç faresi V2 ile karşılaştırıldığında önemli ölçüde daha düşük kod çözme performansı gösterdi (Şekil 1). 6g). Aslında, ağaç faresi V2 aktivitesini kullanarak kod çözme, primat posterior IT’ninkine benzerdi. Beklendiği gibi, primatların ventral görsel hiyerarşisinin zirvesinde yer alan primat anterior IT, en yüksek kod çözme doğruluğunu gösterdi.
Primat ventral akışının ayırt edici özelliği, görüş değişmezliğinin kademeli olarak ortaya çıkmasıdır; bu da benzer bir ilerlemenin ağaç farelerinde var olup olmadığı sorusunu gündeme getirir.8,31,47. DNN modellerini kullanarak, tahmin edilen görünüm değişmezliği indeksini (Yöntemler) 1.593 nesne görüntüsüne verilen yanıtlara dayanmaktadır. Makaklarda, bu tahmin edilen indeks ampirik olarak ölçülen görüş değişmezliği indeksi ile pozitif korelasyona sahipti ve her ikisi de ventral hiyerarşi boyunca arttı (Genişletilmiş Veri Şekil 1). 5a-c). Aynı yaklaşımı ağaç farelerine uyguladığımızda, model tarafından tahmin edilen yanıtlarda böyle bir eğilim bulamadık (Genişletilmiş Veri Şekil 1). 5d, e). Açık bir ilerlemenin olmaması, ağaç sivri faresinin ventral akışında görüş değişmezliğinin aynı hiyerarşik şekilde ortaya çıkmayabileceğini veya mevcut modeller tarafından yakalanamayabileceğini göstermektedir. Bununla birlikte, primatlarda olduğu gibi görüş değişmezliğinin ağaç sivri faresinin görsel yolunun temel düzenleme ilkesi olup olmadığını belirlemek için her alanda doğrudan ampirik testlere ihtiyaç vardır.32 ve kemirgenler48,49. Birlikte ele alındığında, bulgularımız ağaç farelerindeki bir dizi birbirine bağlı alan boyunca görsel işlemenin primatlarla karşılaştırıldığında nasıl olduğunu gösteriyor5,7,8,9,13,32 ve kemirgenler50,51,52hem önemli benzerlikleri hem de farklılıkları vurgulamaktadır (Şekil 1). 6 saat).